? ? ? ?來源:生信人 ? ? ? ?2020年7月6日,美國約翰霍普金斯大學醫學院腫瘤系Sidney Kimmel綜合癌癥研究中心的Jonathan D. Powell教授課題組在Nat Rev Cancer上發表了題為“Metabolism of immune cells in cancer”的綜述 ,試圖闡明癌細胞代謝和免疫代謝之間相互作用。作者認為通過了解包括對癌癥的免疫反應在內的各種細胞的不同代謝需求,可以選擇性調節免疫細胞功能,并對癌癥和免疫新陳代謝的評估,進而揭示新陳代謝的治療窗口并進行干預。 
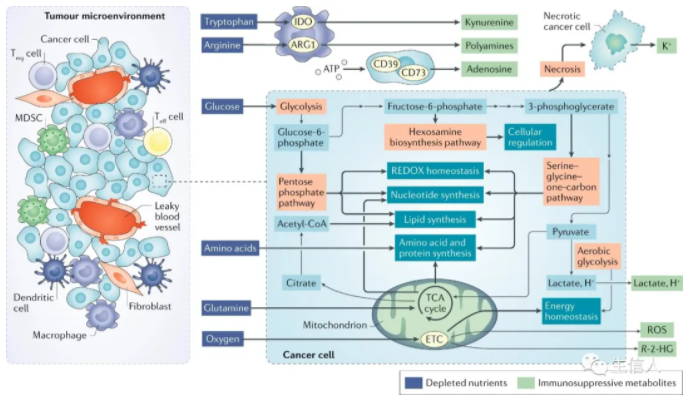
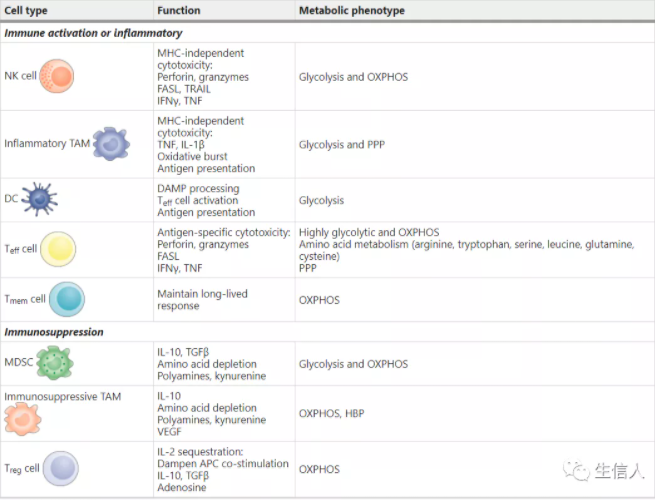
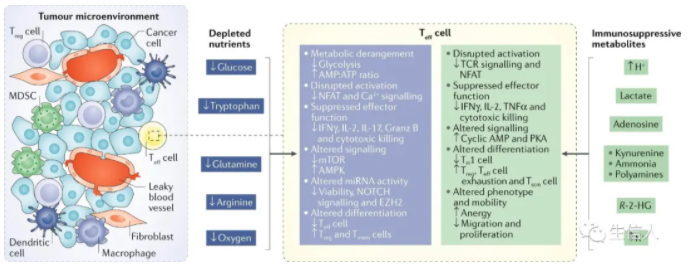
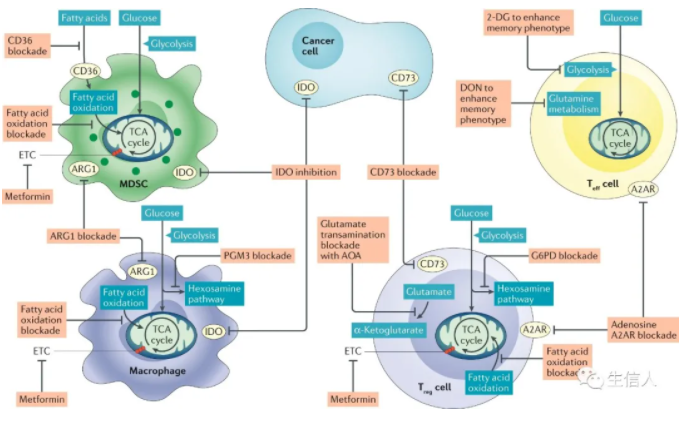
? ? ? ? 摘要 ? ? ? ? 免疫檢查點的封閉和過繼細胞療法的成功,使免疫療法成為治療癌癥的成熟手段。細胞代謝已成為癌細胞和免疫細胞維持活力和功能的關鍵因素。為了維持腫瘤間龐大的合成代謝需求,腫瘤采用了與普通體細胞不同的代謝機制。這種代謝方式可以導致腫瘤微環境呈酸性,低氧,和/或耗盡了免疫細胞所需的關鍵營養物質。在這種情況下,腫瘤代謝本身就可以限制免疫檢查點對腫瘤細胞的破壞。由于我們對免疫細胞代謝和癌癥代謝的理解在過去十年中逐漸加深,我們希望能夠闡明癌細胞代謝和免疫代謝之間相互作用。盡管有些代謝過程似乎是癌癥和免疫細胞應答的基礎,但代謝異質性和可塑性可能有助于區分兩者。因此,了解包括對癌癥的免疫反應在內的各種細胞的不同代謝需求,可以選擇性調節免疫細胞功能。對癌癥和免疫新陳代謝的評估可以揭示新陳代謝的治療窗口并進行干預。 ? ? ? ?前言 ? ? ? ?過去幾十年的研究表明,激活的免疫細胞可以利用多條代謝途徑幫助癌細胞生長(圖1)。這種代謝適應的融合使腫瘤微環境(TME)中的癌細胞和免疫細胞進行營養競爭。然而,我們發現癌細胞與免疫細胞的代謝程序之間以及不同免疫細胞之間存在根本差異。了解這些差異可以為發現針對新陳代謝開展腫瘤治療提供支持。? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? 圖1:TME中的癌細胞代謝和異常 ? ? ? ?癌細胞具有上調葡萄糖的糖酵解分解代謝,形成乳酸鹽的能力,即使在氧氣充足的條件下(有氧糖酵解),該效應被稱為“ Warburg效應”,也被認為是惡性腫瘤的標志。現在人們越來越清楚地認識到,癌癥的代謝也具有異質性,癌細胞可以參與各種代謝程序以滿足生長和增殖的需求,除有氧糖酵解之外,線粒體呼吸也至關重要。我們可以發現具有高代謝活性的癌細胞(圖1)會對TME產生深遠的影響,導致營養物質消耗,缺氧,酸性和代謝產物的產生,這些代謝產物在濃度過高下可能具有毒性。來自TME的大量葡萄糖通過有氧糖酵解代謝,產生大量的乳酸和H +,從而降低了腫瘤內的pH。也就是說,產生乳酸的糖酵解和氧化磷酸化(OXPHOS)之間的平衡可能取決于缺氧的程度,缺氧的程度在TME中既可以是異質的,又可以是廣泛的。值得注意的是,在中度低氧區域,線粒體呼吸產生的CO2被細胞外碳酸酐酶水合,形成HCO3-和H +。因此,氧化代謝可能是TME中細胞外酸化的重要且易被忽視的來源。 ? ? ? ?最近的免疫療法包括免疫檢查點的抑制與過繼細胞療法(CAR T)在腫瘤免疫中得到廣泛應用。例如,黑色素瘤細胞中糖酵解代謝的增加與對過繼性T細胞療法和免疫檢查點的抗性相關。其他研究表明,腫瘤細胞上的免疫檢查點(包括PD1和B7-H3)發出的信號導致TME中的葡萄糖消耗增加。有趣的是,實際上腫瘤酸化的直接結果就是誘導了免疫抑制檢查點相關途徑。此外,免疫檢查點的阻斷可以抑制腫瘤細胞的糖酵解,恢復TME中的葡萄糖,并允許T細胞糖酵解和細胞因子的產生。最近的一些研究表明,針對腫瘤固有代謝的特定方面,例如六胺生物合成途徑(HBP)或谷氨酰胺代謝,可以促進免疫反應并使腫瘤獲得對免疫檢查點封閉的敏感性。 ? ? ? ?由于免疫治療已成為腫瘤治療的支柱,因此深入了解浸潤性免疫細胞與癌癥之間的代謝依賴性變得越來越重要。本綜述旨在討論以下基本問題:哪些代謝程序對于參與癌癥免疫應答的細胞亞群至關重要;這些代謝程序在TME中如何受到干擾;TME中代謝紊亂對當前免疫治療的影響;以及如何利用代謝干預措施來增強抗腫瘤免疫反應。 ? ? ? ?腫瘤免疫微環境 ? ? ? ?癌細胞所特有的高代謝途徑(圖1)對TME中營養成分等影響較大,并可能對免疫反應產生關鍵影響。癌細胞的高代謝活性和TME中紊亂的脈管系統都可以導致營養物質的缺乏和低氧微環境,在癌細胞與浸潤的免疫細胞之間建立代謝競爭。在小鼠模型中,抗腫瘤CD4 + T細胞的葡萄糖攝取和效應功能與癌細胞的糖酵解活性成反比。此外,對來自TCGA的黑色素瘤患者的轉錄組分析顯示,效應T(Teff)細胞基因,例如CD40lg和IFNG,與HK2(糖酵解通路的限速酶)表達呈反相關。在TME細胞內活躍的代謝程序也可能導致某些代謝產物的毒性濃度增加。TME中已報道了腺苷,犬尿氨酸,鳥氨酸,活性氧(ROS)和鉀的水平升高,以及酸中毒增加都會在抑制抗腫瘤免疫反應方面產生深遠影響。 ? ? ? ?TME的免疫環境由一系列不同的細胞類型構成(表1)。效應細胞執行細胞殺傷功能,無論在天然免疫還是適應性免疫反應中。在適應性免疫反應中的抗腫瘤細胞包括CD4 +和CD8 + Teff細胞,它們通過不同機制殺死癌細胞。CD8 + Teff細胞通過誘導凋亡和細胞因子分泌直接殺死腫瘤細胞中。CD4 + T細胞包含許多細胞亞群。其中一些細胞亞群(如Th1)也具有顯著的抗腫瘤活性。這些抗腫瘤CD4 + T細胞(CD4 + conv)與免疫抑制性CD4 + T細胞(Treg)不同。盡管CD4 + conv細胞可能參與直接殺死腫瘤細胞,但它們主要通過細胞因子分泌和協助CD8 + T細胞活化來促進抗腫瘤免疫。抗腫瘤CD4 + conv T細胞與CD8 + Teff細胞具有相同重要的代謝特征。盡管就抗腫瘤免疫性而言,B細胞的研究較少,但B細胞也可能在TME中發揮效應作用。重要的是,作為適應性免疫系統的一部分,T細胞和B細胞會產生記憶細胞群,這些記憶細胞群在感染或腫瘤反應消退后會持續很長時間。CD8 +記憶T細胞(T mem)是長期腫瘤控制的關鍵。先天細胞,例如自然殺傷(NK)細胞和炎性巨噬細胞,也具有關鍵的抗腫瘤效應功能。TME內還有免疫抑制細胞群,包括CD4 + FOXP3 + Treg細胞,髓樣抑制細胞(MDSC),抗炎巨噬細胞和一些B細胞亞群。抗原呈遞細胞,例如腫瘤內樹突狀細胞(DC),在維持TME內的適應性免疫反應起重要作用。 表 1 腫瘤微環境中的細胞組成 ? ? ? ?抗腫瘤反應的代謝 ? ? ? ?抗腫瘤效應T細胞的葡萄糖代謝 ? ? ? ?CD4 + conv和CD8 + Teff細胞形成了抗腫瘤免疫反應的關鍵細胞群。幼稚的T細胞在識別抗原提呈和共刺激因子的信號后,可以進行增殖以及代謝。盡管許多早期研究說明有氧糖酵解的上調是T細胞活化的標志,但現研究表明三羧酸(TCA)循環代謝和OXPHOS上調也是CD4 + conv和CD8 + T細胞活化的關鍵。盡管TCA循環代謝在激活后24 h內被上調,但有氧糖酵解上調更為迅速,發生在激活后6 h內。 ? ? ? ?MYC和缺氧誘導因子1(HIF-1)的轉錄活性均受T細胞活化而上調,并促進代謝重編程。值得注意的是,盡管HIF-1調節缺氧反應的代謝,但在缺氧條件下,其活性也會因響應T細胞活化而被誘導。MYC和HIF-1的轉錄活性導致編碼促進糖酵解的酶(例如丙酮酸激酶(PKM1),己糖激酶2(HK2)和GLUT1)相關基因的表達上調。糖酵解途徑中來自近端代謝產物的途徑也是T細胞活化和功能的組成部分(圖1)。磷酸戊糖途徑(PPP)代謝6-磷酸葡萄糖生成NADPH和5-核糖。在CD4 + T細胞激活后,進入PPP的葡萄糖顯著增加。PPP是NADPH的主要細胞來源,也是新激活的CD8 + T細胞中脂肪酸和質膜合成所必需。NADPH對于REDOX穩態在增殖哺乳動物細胞中也至關重要。激活的T細胞中的ROS水平需要進行精細調節。盡管失調的ROS水平可能具有毒性,但ROS在Teff細胞活化中起著重要作用,已被證明可促進CD4 +和CD8 + T細胞中NFAT依賴性的IL-2表達。另一條起源于早期糖酵解反應的途徑是HBP,是糖基化底物的主要細胞來源,它可介導多種蛋白質,包括穩定性,運輸以及功能作用。HBP依賴于葡萄糖和谷氨酰胺的代謝,并且對它們發生反應。由HBP產生的主要底物UDP-GlcNAc對于效應CD4 +和CD8 + T細胞的擴增和功能至關重要。最后,絲氨酸-甘氨酸-一碳途徑使細胞能夠生成用于葉酸循環的絲氨酸,甘氨酸,NADPH和一碳單位。Teff細胞的增殖和功能在體外和體內都取決于絲氨酸的充分代謝(圖1)。 ? ? ? ?未代謝為乳酸或通過糖酵解途徑代謝的葡萄糖碳對Teff細胞中的TCA循環有重要作用6(圖1)。在高度增殖的細胞中,TCA循環的中間體被迅速消耗,以作為廣泛的生物分子合成的基礎,這一過程稱為 cataplerosis。例如,檸檬酸鹽可以輸出到細胞質中以再生成乙酰輔酶A,用于脂質和膽固醇的合成,這兩者對于在增生的Teff細胞中產生細胞膜至關重要。像癌細胞一樣,Teff細胞具有高度增殖能力,并上調特定的糖酵解程序,包括有氧糖酵解,PPP,HBP和TCA循環,以實現大量的細胞分裂和效應功能。 ? ? ? ?TME中的T細胞和葡萄糖受限 ? ? ? ?TME中的葡萄糖受限會明顯影響T細胞反應。例如,低葡萄糖條件(0.1μmM)抑制了T細胞中糖酵解中間體磷酸烯醇式丙酮酸(PEP)的生成,從而破壞了體外鈣依賴性NFAT信號傳導。與對照相比,降低生長培養基中的葡萄糖濃度可以抑制細胞外酸化率,增加耗氧率,減弱mTOR信號傳導并抑制兩者的效應功能。降低的mTOR復合物1(mTORC1)信號干擾Teff細胞分化,并且在CD4 + T細胞的情況下有利于免疫抑制性Treg細胞的發展。有趣的是,在CD8 + T細胞中,雷帕霉素對mTOR的阻滯促進了Tmem細胞的分化,這可能在維持抗腫瘤反應中發揮重要作用。與對照培養基相比,葡萄糖的下降抑制了Teff細胞中關鍵效應分子干擾素-γ(INFγ),IL-17和顆粒酶B的產生。原發性卵巢癌細胞條件下培養基中的葡萄糖限制導致microRNA介導的組蛋白甲基化酶EZH2的抑制,導致NOTCH信號減少,細胞因子產生受到抑制以及Teff細胞活力降低。 ? ? ? ?在共培養實驗增加小鼠肉瘤細胞的糖酵解能力,可以導致CD8 + T細胞效應功能受到抑制。同樣,與野生型腫瘤相比,在小鼠模型中,植入過表達Hk2的黑色素瘤細胞抑制了CD4 + T細胞的抗腫瘤效應功能和體內反應。此外,黑色素瘤和非小細胞肺癌患者腫瘤樣品中糖酵解相關基因的表達與T細胞浸潤呈負相關。我們也可以通過直接操縱T細胞代謝來實現新陳代謝平衡。例如,與對照相比,在黑色素瘤中腫瘤特異性CD4 + T細胞中糖酵解酶PEP羧激酶的過表達可以改善抗腫瘤反應。 ? ? ? ?線粒體呼吸也是Teff細胞代謝的關鍵。最近的研究報道,癌癥患者的T細胞(與健康對照相比)和荷瘤小鼠的腫瘤浸潤性CD8 + T細胞(與非浸潤性CD8 + T細胞相比)顯示線粒體質量降低以及線粒體功能障礙。與健康對照相比,慢性淋巴細胞性白血病患者外周CD8 + T細胞的線粒體受損。此外,這些患者對CAR T細胞療法的反應程度與注入的CAR T細胞的線粒體損傷程度呈負相關。與健康供體的CD8 + T細胞相比,腎細胞癌患者的腫瘤浸潤性CD8 + T細胞顯示線粒體動力學和功能失調,包括線粒體ROS水平升高和超極化。這些T細胞的正常體外活化可以通過線粒體ROS清除劑或丙酮酸補充進行挽救。線粒體的生物發生和功能在功能失調的腫瘤浸潤性CD8 + T細胞的亞群(特別是耗竭性T細胞)中極為紊亂。總體而言,這些研究表明癌癥本身可以導致Teff細胞代謝紊亂,包括線粒體動力學,并且癌細胞的糖酵解活性程度與浸潤性T細胞的抗腫瘤效應功能之間存在相互關系。 ? ? ? ?氨基酸與抗腫瘤T細胞反應 ? ? ? ?像癌細胞一樣,高度增殖的免疫細胞(例如活化的T細胞)也依賴于氨基酸代謝來支持蛋白質和核苷酸的合成。因此,氨基酸轉運蛋白,包括SLC7A5(也稱為LAT1),SLC38A1(也稱為SNAT1),SLC38A2(也稱為SNAT2)和SLC1A5(也稱為ASCT2)在T細胞激活后被高度上調。必需氨基酸必須是外源獲得。例如,亮氨酸是效應CD8 +和CD4 + conv T細胞中mTORC1信號傳導,效應功能和分化所必需。活化的T細胞還可以快速代謝精氨酸,外源性精氨酸的補充可以改善T細胞的適應性并增加中央Tmem細胞的生成。絲氨酸,色氨酸和半胱氨酸也是T細胞反應的重要營養成分,因此是抗腫瘤免疫反應的重要介質。色氨酸是必需氨基酸,在TME中是決定T細胞反應強度和有效性的重要因素。與正常培養基相比,在不含色氨酸的培養基中,人類T細胞的增殖和活化受到了強烈抑制。癌細胞,腫瘤相關巨噬細胞(TAM),MDSC,抑制性DC和癌癥相關成纖維細胞可通過吲哚胺2,3-二加氧酶(IDO)的酶促活性耗盡色氨酸水平,這些酶可在TME內高水平表達。代謝途徑對腫瘤生長的十分重要,IDO表達與多種癌癥類型(包括胃癌,結腸直腸癌,非小細胞肺癌和黑色素瘤)患者的不良預后相關。 ? ? ? ?在增殖細胞中,谷氨酰胺為氨基酸和核酸的合成提供氮,為TCA循環中間體提供補充的碳,這個過程被稱為“ anaplerosis”(圖1)。癌細胞和一些活化的免疫細胞,例如T細胞和巨噬細胞,通常富含高度谷氨酰胺。在體外刺激鼠CD4 + conv T細胞的過程中,谷氨酰胺轉運蛋白SLC1A5和SLC38A1和/或SLC38A2的表達顯著上調。在MYC的驅動下,谷氨酰胺被谷氨酰胺酶(GLS)代謝為谷氨酸,在由谷氨酸脫氫酶(GLUD1)轉化為α-酮戊二酸(αKG;也稱為2-氧戊二酸)后,谷氨酰胺可能進入TCA循環。隨后在TCA周期中,αKG被代謝為琥珀酸鹽和富馬酸鹽。有趣的是,在谷氨酰胺受限的情況下,一些癌細胞系轉換為葡萄糖引起的 anaplerosis,其中丙酮酸被丙酮酸羧化酶轉化為草酰乙酸,進入TCA循環。最近表明,在體外谷氨酰胺阻斷的條件下,效應CD8 + T細胞也能夠上調丙酮酸羧化酶的活性。 ? ? ? ?盡管通過限制培養基中的谷氨酰胺可以抑制分化的CD8 + Teff細胞的效應功能和增殖,但如果在激活CD8 + T細胞的過程中限制了谷氨酰胺的利用率,它會改變向CD8+記憶表型的分化,研究表明分化作用是由αKG介導的。αKG和其他TCA代謝物(例如琥珀酸酯和富馬酸酯)可以調節多種細胞過程的活性,包括表觀遺傳重塑和關鍵轉錄因子(如HIF-1α)的穩定性。 ? ? ? ?脂質代謝與T細胞 ? ? ? ?活化的T細胞還可以重新編程脂質代謝,上調脂質合成和膽固醇攝取,其對細胞膜合成至關重要,并分別由轉錄因子固醇調節元件結合蛋白1(SREBP1)和SREBP2介導。在缺少SREBP1和SREBP2功能的活化小鼠CD8 + T細胞中,增殖,代謝重編程和抗病毒活性被顯著抑制。此外,在體外CD8 + T細胞活化和擴增過程中,細胞膜中的膽固醇含量部分受膽固醇酯化酶乙酰輔酶A乙酰轉移酶(ACAT1)調節。在過繼轉移小鼠腫瘤模型中,Acat1基因敲除的CD8 + T細胞表現出增加的膜膽固醇和改善的T細胞受體聚集和信號傳導,從而導致增殖,功能增強和殺傷力增強。 ? ? ? ?用阿瓦西米對ACAT1的藥理抑制作用改善了小鼠的抗腫瘤作用。最新研究 證明腫瘤中高膽固醇可通過激活內質網應激反應而誘發T細胞功能障礙。因此,盡管膽固醇對于Teff細胞的增殖和代謝很重要,但針對膽固醇代謝特定方面改善抗腫瘤免疫反應的益處尚需進一步研究。 ? ? ? ?免疫記憶的代謝 ? ? ? ?與Teff細胞不同,CD8 + Tmem細胞優先依賴OXPHOS。與CD8 + Teff細胞相比,增強的備用呼吸能力也是Tmem細胞的高度特征。最初研究顯示使用埃托莫西酯作為肉堿棕櫚酰轉移酶1A(CPT1A)的抑制劑,該酶是線粒體轉運蛋白,負責長鏈脂肪酸的進入以進行脂肪酸β-氧化(FAO),表明FAO是Tmem中OXPHOS的主要燃料。然而,最近使用T細胞特異性Cpt1a敲除模型進行的研究對此提出了質疑,并證明了高劑量依托莫昔(200μm)的脫靶效應可能是原因。 ? ? ? ?TCA循環的中間體,例如αKG,琥珀酸酯和富馬酸酯,在適應性記憶中特別重要。通過改變這些TCA代謝產物抑制2-氧戊二酸依賴性雙加氧酶(2OGDD)已顯示可增加CD8 + T細胞中的記憶細胞分化。盡管葡萄糖,谷氨酰胺和脂肪酸是推動TCA循環的主要營養來源,但一系列其他營養物質(例如氨基酸和乙酸鹽)也可以進入該循環。特別是,乙酸酯代謝正在成為CD8 + T細胞和某些癌癥類型中乙酰輔酶A的重要來源。在線粒體中,乙酸鹽在被酰基輔酶A合成酶短鏈家族成員1(ACSS1)代謝形成乙酰輔酶A后可以進入TCA循環。另外,乙酸鹽可以在細胞質中通過ACSS2轉化為乙酰輔酶A,在乙酸中它可以促進脂肪酸合成和乙酰化反應,這對于表觀遺傳重編程和翻譯后修飾有重要意義。乙酸鹽的代謝是促進記憶CD8 + T細胞功能的重要代謝途徑。有趣的是,在T細胞活化過程中,谷氨酰胺代謝的阻滯增加了Tmem細胞的分化,并誘導了乙酸代謝和相關的酶,包括ACSS1和ACSS216,85。作為靜止細胞,Tmem細胞相對于有氧糖酵解優先依賴于OXPHOS,并且具有顯著的線粒體儲備,這是抗原活化后進一步上調OXPHOS所必需。Tmem細胞可以采用幾種不同的營養來源,以促進這種代謝程序。 ? ? ? ?缺氧與抗腫瘤T細胞反應 ? ? ? ?盡管腫瘤具有高度異質性,但高水平的代謝活性和相關的氧氣消耗以及功能紊亂,功能差的脈管系統仍會產生缺氧區域,中位氧飽和度水平低于2%(正常組織中值約為5%) 。缺氧對Teff細胞的影響并不直接。在常氧條件下,HIF-1轉錄活性會響應T細胞活化而被上調,因此要了解缺氧對進一步增強HIF-1活性的影響,同時還要評估HIF-1的依賴效應,具有一定的挑戰性。早期對CD8 + Teff細胞活化,分化和功能的體外研究表明,盡管在缺氧條件下抑制了增殖和某些細胞因子的表達,但其溶解能力,活化標記物和存活率卻得到了改善。隨后的體內研究表明,與暴露于環境O2張力(20%)的小鼠相比,暴露于低于大氣O2張力(8%)的小鼠中CD4 +和CD8 +脾T細胞的活化更弱。其他研究表明,體外低氧暴露會導致代謝物(S)-2-羥基戊二酸(S-2-HG)在細胞內積聚,從而深刻改變CD8 + T細胞的活化和分化,抑制細胞因子的分泌和溶細胞能力。但有趣的是,在小鼠模型體內轉移后增強增殖,長期生存和抗腫瘤反應。糖酵解活性通過組成性HIF-1α活性(通過有條件地敲除HIF-1調節劑Vhl來實現)實際上促進了疫苗模型中長效效應記憶細胞的數量。其他工作表明,缺氧誘導了TME中各種細胞外切核苷酸酶CD39和CD73的表達,這些酶將TME中的ATP分解為腺苷。腺苷是A2A和A2B嘌呤能受體的配體,它們在廣泛的免疫細胞中表達,并且廣泛地具有免疫抑制作用,抑制效應細胞功能和Teff細胞增殖。有趣的是,補充氧氣通過下調腺苷信號傳導途徑來增強小鼠T細胞的抗腫瘤免疫反應。缺氧對抗腫瘤T細胞的影響是一個不斷發展的研究領域。考慮到腫瘤中低氧區域的普及以及低氧對適應性免疫反應的深遠影響,其進一步的研究將顯然有益于免疫治療領域。 ? ? ? ?毒性代謝物 ? ? ? ?除腺苷外,癌細胞代謝產生的許多其他產物也會影響浸潤的T細胞(圖1和2)。在小鼠和人CD8 + T細胞的體外研究中,TME中細胞外乳酸和H +的水平升高可以抑制T細胞增殖,存活,細胞毒性和細胞因子的產生。與標準生長培養基相比,在高水平的乳酸和H +存在下,小鼠CD8 + T細胞的體外激活過程中,編碼關鍵Teff細胞轉錄因子NFAT的基因的上調受到損害。體內小鼠研究表明,與空載細胞相比,被Ldha敲除的小鼠黑色素瘤細胞產生的乳酸更少,對免疫介導的腫瘤排斥反應的反應更強。與對照培養基相比,在乳酸和H +升高的情況下激活的人效應CD8 + T細胞中,MAP激酶信號傳導也受到嚴重破壞。? ? ? ? ? ? 圖2:TME中的代謝紊亂抑制T細胞功能 ? ? ? ?腫瘤中特定氨基酸的積累也可以抑制Teff細胞反應。在這方面,研究得最好的是色氨酸代謝產物,尤其是犬尿氨酸,其作用是通過IDO1的活性產生。與對照相比,犬尿氨酸在體外引起活化CD8 + T細胞上PD1協同抑制途徑的上調。與對照相比,在用外源性犬尿氨酸處理的小鼠模型中,在腫瘤浸潤的CD8 + T細胞上也觀察到PD1的這種上調。來自經犬尿氨酸處理的荷瘤小鼠的腫瘤浸潤CD8 + T細胞產生的IFNγ和TNF較少。 ? ? ? ?在報道說明癌細胞通過釋放競爭性代謝物(R)-2-羥基戊二酸酯(R-2-HG)抑制T細胞活性。這種代謝物可以抑制表觀遺傳的雙加氧酶,例如組蛋白脫甲基酶,從而導致甲基化增強和轉錄修飾。由異檸檬酸脫氫酶(IDH)突變的人腦膠質瘤產生的R-2-HG在體外研究中被T細胞吸收。R-2-HG在體外干擾活化CD4 +和CD8 + T細胞的增殖,T細胞受體信號傳導,NFAT活性和多胺生物合成。在IDH突變的神經膠質瘤小鼠中R-2-HG釋放入TME抑制補體介導的抗腫瘤反應以及T細胞遷移,增殖和細胞因子分泌。這些研究突出了TME內癌癥代謝物和免疫功能之間的復雜相互作用(圖2)。 ? ? ? 高度壞死導致TME中鉀的含量增加,從而限制了T細胞效應功能。該狀態由降低的乙酰輔酶A胞質水平介導,此狀態誘導了活化T細胞的表觀遺傳重塑,從而導致TME中Teff細胞功能異常。但是,這種功能障礙狀態富含T細胞干性的特征。高鉀濃度的Teff細胞在離體刺激和擴增后產生的T細胞具有更高的體內持久性,多能性和腫瘤清除能力。通過腫瘤壞死和TME固有的代謝活性產生T細胞抑制性代謝產物,是逃避腫瘤免疫的重要機制。 ? ? ? ?代謝和天然效應反應 ? ? ? ?由于NK細胞在主要組織相容性復合物I類(MHC-I)下調(癌細胞的常見逃避策略)期間殺死細胞,所以它們形成了先天反應的關鍵效應組成成分。在體外,NK細胞的細胞因子刺激(IL-12和IL-15)后,有氧糖酵解和OXPHOS在代謝上被上調。有趣的是,在體外NK細胞刺激過程中,這些細胞因子誘導的代謝變化需要SREBP轉錄因子。在體外,SREBP活性的藥理抑制作用抑制了代謝重編程,細胞因子產生和細胞毒性,并在過繼性NK細胞小鼠模型中減少了抗腫瘤反應。有趣的是,據報道內源性SREBP抑制劑(例如27-羥基膽固醇)可在TME中增加,因此可能是NK細胞抑制的機制。小鼠的肺癌進展和與腫瘤相關的轉化生長因子-β(TGFβ)與腫瘤相關的NK細胞中果糖-1,6-雙磷酸酶(FBP1)表達的增加有關。FBP1是糖異生的關鍵酶,一旦激活,FBP1會強烈抑制NK細胞的糖酵解,從而導致功能障礙和活力降低。有趣的是,FBP1的藥理抑制作用足以在體外重建糖酵解代謝以及細胞因子的產生和細胞毒性,并改善過繼細胞療法小鼠模型的抗腫瘤反應。這些研究表明,通過FBP1抑制來挽救NK功能取決于葡萄糖代謝的恢復,因為用2-脫氧葡萄糖(2-DG)阻斷葡萄糖代謝阻止了FBP1抑制引起的拯救。2-DG本身也導致NK細胞功能異常,這表明抑制葡萄糖代謝可能對NK細胞抗腫瘤反應產生深遠影響。TME內的其他代謝紊亂也可能影響NK細胞功能。例如,精氨酸水平低會損害NK細胞的增殖和IFNγ的產生,而缺氧會抑制細胞溶解活性。在體外研究中,人類NK細胞激活受體(例如NKp46和NKp30)在響應低氧或精氨酸水平較低時受到抑制。如在TME中一樣,高乳酸水平和相關的低pH值在體外研究中也抑制了NK細胞的細胞毒性,細胞因子產生和NFAT信號傳導。最后,TME中腺苷水平升高可強烈抑制NK細胞效應功能和增殖。 ? ? ? ?其他先天細胞,巨噬細胞和DC也會在激活時引發特定的代謝程序。在早期的體外研究中,將巨噬細胞分為炎性(M1)或免疫抑制(M2)表型,但很少有證據表明這些極化表型在體內發揮著獨特的作用。最近發現了以不同轉錄狀態為特征的一系列巨噬細胞表型。也就是說,具有炎性特征的巨噬細胞在抗腫瘤免疫中發揮重要作用。葡萄糖代謝是巨噬細胞中炎癥表型的重要方面。被Toll樣受體激動劑激活后,這些細胞顯示出糖酵解基因的表達增加,葡萄糖攝取的高水平,乳酸的產生增加以及谷氨酰胺過表達。這種代謝重編程導致琥珀酸水平升高,從而通過穩定HIF-1而增加了炎性細胞因子IL-1β的表達。炎性巨噬細胞還特別依賴于PPP來產生NADPH,13C-葡萄糖示蹤研究證實,通過激活培養物中的炎性表型,通過該途徑的葡萄糖路徑增加。NADPH對于產生高水平的ROS(作為氧化性爆發的一部分)是必需的,這是這些細胞的關鍵效應機制。精氨酸也是促炎性“ M1”巨噬細胞功能中的關鍵營養物質,因為在體外研究中,精氨酸比其他活化或“ M2”極化巨噬細胞表達更高水平的誘導型一氧化氮合酶(iNOS)。iNOS需要精氨酸才能產生細胞毒性一氧化氮,這是抗腫瘤反應的重要促炎介質。 ? ? ? ?TME中特定的營養缺乏,特別是葡萄糖和精氨酸,會嚴重限制這些細胞中新陳代謝和效應程序。葡萄糖限制不僅可以整體抑制糖酵解,而且可以降低PPP活性和TCA循環功能,從而限制了NADPH,ROS和琥珀酸酯的生成,所有這些都可以嚴重限制M1巨噬細胞的功能。2-DG的糖酵解抑制作用顯著減少了巨噬細胞促炎細胞因子的分泌。 ? ? ? ?DC是參與抗腫瘤應答的一類重要的抗原呈遞細胞,能夠進行抗原交叉呈遞的腫瘤內DC已作為這種反應的重要組成部分。激活后,DC會成熟并允許抗原加工并呈遞給T細胞。該反應與從OXPHOS到有氧糖酵解的代謝轉換相關,其由HIF-1α響應體外LPS激活,并由PI3K-AKT途徑響應Toll樣受體體外刺激介導。在DC激活過程中,這種轉換為糖酵解作用并遠離OXPHOS的過程對于DC存活,刺激性細胞因子的產生以及T細胞的激活至關重要。有趣的是,AMPK的藥理學激活可促進線粒體的生物發生和氧化呼吸,并足以在體外阻止DC成熟。鑒于這種對有氧糖酵解的關鍵依賴性,TME中的葡萄糖競爭可能會顯著抑制DC激活和生存能力,從而限制DC促進有效和持久性T細胞反應的能力。 ? ? ? ?癌癥免疫逃逸的新陳代謝 ? ? ? ?適應性免疫抑制的代謝 ? ? ? ?免疫抑制性Treg細胞優先依賴TCA循環和線粒體呼吸。盡管初步研究表明Treg細胞對FAO的依賴性并未說明依托莫昔的脫靶作用,但其他研究表明,FAO確實支持Treg細胞中的OXPHOS,盡管不是唯一途徑。與Teff細胞相反,Treg細胞在體外顯示降低葡萄糖攝取并表達較低水平的GLUT1的表達。有趣的是,盡管糖酵解似乎并未在Treg細胞分化或長壽表型中發揮關鍵作用,但據我們發現,一部分高活性Treg細胞(稱為效應Treg細胞)依賴于糖酵解的上調以獲得最佳功能。Treg細胞似乎具有新陳代謝的靈活性,可以在相對苛刻和異質的條件下(例如TME)生長。為此,據報道,Treg細胞轉錄因子FOXP3通過抑制MYC(有利于OXPHOS和NAD(H)氧化)來重新編程細胞代謝。在低葡萄糖和高乳酸的條件下(例如在TME中發現),這些適應作用使這些免疫抑制細胞具有代謝優勢,從而使Treg細胞能夠在體外抵抗乳酸誘導的功能性和增殖性抑制(不同于Teff細胞)。在體外實驗中,培養基中的葡萄糖或谷氨酰胺缺乏(導致細胞內αKG減少)可以改變CD4分化并促進Treg細胞的發育。 ? ? ? ?與Teff細胞相似,Treg細胞對缺氧的反應尚不完全清楚。缺氧已顯示可促進細胞因子介導的Treg細胞募集進入腫瘤環境。FOXP3轉錄本實際上是響應HIF-1α誘導而增加。此外,與野生型對照相比,過繼轉移的Treg細胞特異性Hif1敲除細胞在小鼠模型中未能遷移到腦腫瘤中,在用二氯乙酸酯處理的Treg細胞中也觀察到糖酵解受到抑制。有趣的是,在混合淋巴細胞培養實驗中,通過Treg細胞上腺苷受體A2A的低氧反應性腺苷信號傳導誘導了增殖,并顯著增強了免疫調節活性。有文獻報道低氧誘導的HIF-1α可以破壞Treg細胞的穩定性,并表明低氧可以通過直接HIF-1α與分別定義細胞亞型的轉錄因子FOXP3和RORγt的直接相互作用來促進TH17 CD4 + T細胞。 ? ? ? ?TME中氨基酸的獨特代謝也可以對Treg細胞產生深遠的影響。IDO1活性可以在體外強烈促進Treg細胞分化,這種作用似乎是色氨酸缺乏癥以及下游代謝產物(如犬尿氨酸)的產生所致。 ? ? ? ?天然免疫抑制的代謝 ? ? ? ?TAM表型為高度免疫抑制。像Treg細胞一樣,M2巨噬細胞上調FAO和線粒體呼吸。盡管早期研究表明FAO必須在M2巨噬細胞中發揮作用,但并未考慮到CPT1A抑制劑etomoxir的脫靶作用,但由于過表達Pgc1α引發巨噬細胞而導致免疫抑制表型,并且抑制促炎細胞因子的產生。M2巨噬細胞以不同于炎性巨噬細胞的方式代謝氨基酸,表達高水平的精氨酸酶1(ARG1),這會消耗精氨酸并生成多胺,多胺是傷口愈合的重要介質,但也具有高度免疫抑制作用。 ? ? ? ?另一組與腫瘤相關的免疫抑制先天細胞MDSC似乎具有高代謝活性。與周圍的MDSCs相比,與腫瘤相關的MDSCs中有氧糖酵解和OXPHOS均上調。在另一項研究中,與來自相同小鼠的脾嗜中性白細胞相比,來自荷瘤小鼠脾臟的粒細胞MDSCs也顯示出有氧糖酵解和OXPHOS的增加。有趣的是,在小鼠乳腺癌模型中,MDSC的體外擴增和在TME中的蓄積可通過用2-DG阻斷糖酵解來減弱,這很可能是由于這些細胞中的ROS水平升高所致。 ? ? ? ?腫瘤中的缺氧區域與巨噬細胞的積累有關,它們在巨噬細胞中通過產生與腫瘤轉移有關的血管生成因子,促有絲分裂因子和細胞因子來協助腫瘤的發展。此外,缺氧可促進免疫抑制巨噬細胞表型的產生。缺氧可產生腺苷,可通過巨噬細胞上的A2A和A2B受體觸發信號傳導,這兩者均可增強免疫抑制巨噬細胞的分化和功能能力,并減弱體外炎性巨噬細胞的細胞因子釋放。已證明培養物中乳酸的升高可促進M2表型,增加ARG1表達和多胺依賴性免疫抑制。在三陰性乳腺癌中,較高的糖酵解率顯示出促進MDSCs,而在這些癌細胞中限制糖酵解則抑制了癌細胞中粒細胞集落刺激因子和粒細胞-巨噬細胞集落刺激因子的分泌,并限制了MDSC的發育。有趣的是,與常氧性MDSCs相比,低氧使MDSCs趨向于具有免疫抑制性的M2樣TAM表型。這是通過HIF-1α機制引發,因為在小鼠黑素瘤腫瘤模型中,與野生型MDSC相比,Hif1a基因敲除的MDSCs顯示出腫瘤生長增加。 ? ? ? ?利用不同代謝的可塑性 ? ? ? ?雖然可以通過抑制許多代謝途徑來減弱Teff細胞的激活,增殖和功能,但是其他屬性,例如重新刺激時的長期生存力或效應功能可能增強。盡管用2-DG抑制糖酵解代謝抑制了Teff細胞的產生,但它也使T細胞趨向于長效記憶表型。有趣的是,在再次輸注用于腫瘤治療之前,在離體T細胞活化和擴增過程中阻斷糖酵解,不僅可以提高抗腫瘤T細胞的存活率,還可以改善細胞因子的產生和細胞毒性。此外,有研究證明了該途徑在維持谷氨酰胺阻斷的CD8 + T細胞中維持代謝穩態方面的重要性。這些發現可能表明一種普遍的治療策略,即阻止使用典型的代謝燃料(例如葡萄糖或谷氨酰胺)可能使某些癌癥在代謝上受到損害,但鑒于抗腫瘤性T細胞使用其他來源的能力,它們可能會在代謝方面保持完整和正常運轉,如醋酸鹽。盡管可以通過藥理學方法將特定的代謝干預作為檢查點封閉的輔助手段引入,但這些靶點可能特別適用于CAR T細胞療法,其中可以通過遺傳手段精確定義代謝途徑。 ? ? ? ?免疫檢查點封閉與免疫代謝 ? ? ? ?定義檢查點療法的代謝后果和其反應后的代謝情況都非常有意義。在多項研究中,檢查點信號已顯示出調節代謝的作用。例如,癌細胞上PDL1的表達可以驅動癌細胞中Akt-mTOR的活化和糖酵解,增加葡萄糖的攝取并增強與T細胞競爭葡萄糖的能力。人胃癌組織中T細胞中的CD155-TIGIT信號傳導抑制了葡萄糖的攝取,乳酸的產生以及糖酵解酶GLUT1和HK2180的表達。相反,與同種型處理的對照T細胞相比,共刺激途徑GITR的激動作用廣泛提高了T細胞的代謝活性和增殖。最后,活化的人T細胞上的體外PD1和CTLA4信號抑制了與T細胞活化相關的代謝途徑,如有氧糖酵解。為此,將代謝抑制劑與檢查點抑制劑結合使用的前景有望提高檢查點封閉的療效。同樣,通過對T細胞進行代謝重編程以使其更強大,持久的記憶細胞可能會改善其對檢查點抑制劑的反應。最近的臨床試驗預示了抗葉酸培美曲塞與抗PDL1免疫檢查點封閉的結合。除具有直接的抗腫瘤作用外,培美曲塞治療還增強了抗腫瘤CD8 + T細胞的代謝適應性和效應功能,并誘導了癌細胞的免疫細胞死亡,從而觸發了免疫反應。 圖3:增強癌癥免疫反應的潛在代謝靶點 ? ? ? ?結論與展望 ? ? ? ?盡管免疫代謝的許多基礎已從癌癥代謝的觀察中獲悉,但很明顯,癌癥與免疫代謝重編程之間存在明顯差異。這些差異為靶向代謝提供機會,以此作為增強免疫療法功效的手段(圖3)。這種方法可以通過許多不同的策略來實現。這些措施包括靶向腫瘤代謝程序以抑制生長和改變TME,靶向抑制性免疫細胞的代謝以抑制其功能以及靶向效應細胞代謝以增強腫瘤殺傷力。同樣,在過繼細胞治療之前,T細胞代謝途徑的體外藥理或基因重編程也可以增強其功能,包括延長壽命或增強效應功能。 ? ? ? ?未來工作應集中于TME內免疫細胞和癌細胞的代謝相互依賴性。除了營養消耗和可以抑制一定濃度下的免疫反應的代謝產物產生之外,癌細胞還可以與TME中的其他細胞發生代謝擾動,其中可以誘導并共同的代謝程序,從而有利于惡性進展。據報道,胰腺星狀細胞可以為癌細胞提供丙氨酸,并且已經報道了骨髓基質細胞提供半胱氨酸以促進慢性淋巴細胞性白血病細胞的存活。在另一份報告中,來自癌細胞谷氨酰胺代謝的氨通過TME擴散并觸發了與癌癥相關的成纖維細胞的自噬,這反過來又提供了蛋白質分解產物,例如谷氨酰胺本身,以進一步支持癌細胞代謝。重要的是要了解逃避免疫的癌癥是否以及通過何種機制選擇免疫細胞的代謝機制,并從其顯著的代謝靈活性中受益。 ? ? ? ?如涉及知識產權請與我司聯系
|