來源:Biotech VentereCapital ?
1.?介紹?
生物技術的進步導致抗體生產和重組技術的改進,推動了抗體和無數抗體結構的開發。目前,美國食品和藥物管理局(FDA)批準了72種抗體,其中30種已注冊用于治療癌癥患者。抗體在癌癥治療中發揮著越來越重要的作用。對于抗體以及如何改變它們的藥代動力學和物理化學性質的理解不斷深入。在抗體作為標準治療方法建立后,越來越復雜的抗體結構被開發出來。除了完整的免疫球蛋白G (IgG)抗體、首個抗體藥物偶聯物和雙特異性抗體(bsAb)已被批準用于治療癌癥患者,其他一些抗體結構藥物正處于臨床試驗中(圖1)。?
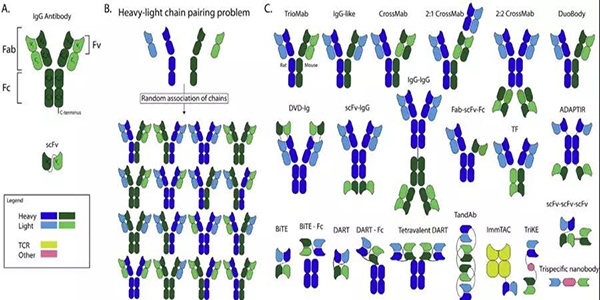
?圖1.目前在臨床試驗中評估的抗體結構和bsAb結構示意圖。(A)IgG抗體結構由Fab和Fc區組成。該Fab區的結合部分稱為單鏈可變片段(scFv)。抗體存在兩條重鏈(VH和CH)和兩條輕鏈(VL和CL)。這些重輕鏈可以通過可變區(VH和VL)和恒定區(CH和CL)進行細分。(B)隨機重-?輕鏈配對可能產生的不同組合。(C)目前已批準或正在臨床試驗中的雙特異性抗體結構
標準的人體抗體是一種專一性抗體,它的兩個結合位點都針對同一靶標。bsAb是一個更為復雜的結構,其結合位點針對不同的作用靶標。這使其能夠實現一些新穎獨特的作用機制,如使免疫細胞參與殺傷腫瘤細胞,向腫瘤細胞遞送有效載荷,或者阻斷腫瘤細胞重要的信號通路(圖2)。每一種作用機制都需要通過修飾bsAb獲得理想的藥代動力學特性。關于這些bsAb結構及其作用機制已經發表了大量的臨床前研究數據。
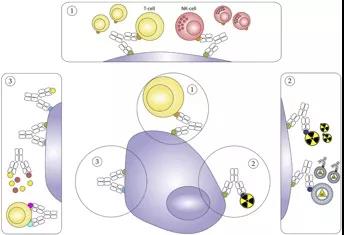 ? ?
圖2.在腫瘤學臨床試驗中提出的雙特異性抗體(bsAbs)作用機制的簡化示意圖。1.免疫細胞與腫瘤細胞的結合。免疫細胞可以與腫瘤細胞結合。2.靶向性遞送有效載荷。用對腫瘤和有效載荷都具有親和力的bsAb靶向腫瘤細胞。3.阻斷信號。bsAb阻斷兩個信號。
在腫瘤學領域,已有兩種雙特異性抗體(bsAbs)被批準用于臨床。Catumaxomab靶向上皮細胞黏附分子(EpCAM)和CD3, 2009年被歐洲藥物管理局(EMA)批準用于治療惡性腹水。然而,應MAH的要求,Catumaxomab的市場授權于2017年6月撤銷。Blinatumomab是靶向CD19和CD3的雙特異性抗體,于2014年12月獲FDA批準,2015年12月獲EMA批準用于治療費城染色體陰性B細胞急性淋巴細胞白血病(ALL) 。在腫瘤之外,結合凝血因子IXa和X的bsAb emicizumab于2017年11月被FDA批準,并于2018年3月被EMA批準用于治療血友病A。
目前,包括blinatumomab在內的57種bsAbs正在用于癌癥患者的臨床試驗中,其中38種使用相同的作用機制:促進免疫細胞與腫瘤細胞的結合。在剩下的19個進行臨床試驗的bsAbs中,5個雙特異抗體的作用機制是向腫瘤遞送有效載荷,14個雙特異性抗體的作用機制是阻斷癌癥微環境中的信號通路。
本綜述的目的有兩個:1)通過總結bsAbs的腫瘤臨床開發來評估雙特異抗體的結構選擇;
? ? ? ? ? ? ? ? ? ? ? ? ? ? ?? 2)明確雙特異性抗體在腫瘤臨床開發中所面臨的挑戰。
??
2.生產雙特異性抗體
單抗的兩個結合區域針對同一表位。因此,單抗是雙價且具有靶標單一性。相反,bsAbs具有能結合兩個不同的靶標的表位,根據其結構的不同,可以是單價的,也可以是雙價的。
單抗通常由雜交瘤細胞系表達,雜交瘤細胞系是分泌抗體的B細胞和永生的骨髓瘤細胞系的融合體。BsAbs可以通過融合兩個雜交瘤細胞系而產生,其結果是產生IgG分子的混合物。此外雙抗還可以通過組合兩種現有抗體或它們的片段來產生。另一種因其靈活性而廣受歡迎的選擇是使用重組蛋白。使用遺傳工程化重組蛋白質創造了關于起源組成和生產系統。例如,這些蛋白質可以用來控制重鏈和輕鏈的結合。基本的bsAb結構包括來自一種抗體的重輕鏈對和來自另一種抗體的重輕鏈對。當這四個單獨的鏈結合在一起時,它們隨機結合,可以產生16種IgG分子的組合。其中的兩種組合產生了所需的bsAbs結構,其異質二聚重鏈與來自同一抗體的特定輕鏈結合(圖1B)。嵌合四聚體、常見的輕鏈和重組蛋白可以通過限制締合的選擇來提供解決方案。嵌合四聚體具有受物種限制的重-輕鏈配對。此外,使用普通的輕鏈也可以防止不必要的重輕鏈組合。重組蛋白可以通過多種途徑促進重輕鏈與重鏈的正確結合。
例如,孔內旋鈕方法,其中一個重鏈是由一個由相對較大的氨基酸組成的旋鈕設計的,而另一個重鏈是由一個由相對較小的氨基酸組成的孔設計的。其他的例子是由肽鏈連接片段的結構,例如雙特異性T細胞銜接器(BiTE)分子,從而繞過鏈的隨機關聯。?
?
?3.合理的設計
就像抗體一樣,bsAb可以通過無數種方式進行修飾,自定義其功能,增強其功效,如調節抗體的免疫原性、效應函數和半衰期。
關于調節免疫原性的問題,抗體結構中來源于小鼠的能產生免疫原性的結構常常被人類的相同結構替代,以降低自身免疫原性。這導致了嵌合人源化抗體結構的產生。越來越多的全人源化抗體生產出來,通常是通過噬菌體展示或產生人類IgG的轉基因小鼠。利用噬菌體展示,篩選表達抗體部分的噬菌體文庫,以確定其與抗原的親和力。其他能夠引起免疫原性的抗體結構的部分是外來氨基酸序列,可能是由新的蛋白質工程引入的。
關于抗體的效應功能,Fc區域在介導這一過程中起著重要作用。該區域參與免疫介導的細胞殺傷機制,如補體依賴的細胞毒性和抗體依賴的細胞毒性。與靶向腫瘤細胞的抗體不同的是,結合免疫細胞的抗體被設計用來減輕這種細胞殺傷作用。腫瘤細胞靶向抗體需要一個功能Fc區域來殺死目標細胞。
糖工程和改變Fc區氨基酸序列可以影響免疫介導的細胞殺傷機制。這些技術可以通過抗體增強或減少免疫介導的細胞殺傷,這取決于修飾抗體的聚糖和氨基酸的位置和功能。除了消除通過免疫介導的細胞殺傷,整個Fc區域也可以被刪除,從而區分出帶有Fc區域的抗體和缺乏Fc區域的抗體。這種去除Fc的抗體的分子量大大降低,進而影響了抗體的藥物動力學特性,包括抗體清除和腫瘤穿透性。
完整的IgG抗體大小約為150 kDa,由肝臟清除,而分子量低于60 kDa的蛋白質則由腎臟清除。腎清除比肝清除快。抗體的大小也可以通過移除Fab中非結合區域CL和CH1區域。如果從結構中刪除非結合區域,則只保留必要的結合區域,即可變區片段。這些由一個單一的肽鏈連接在一起的可變區片段稱為單鏈可變片段(scFv) 。由于ScFvs體積小且缺乏新生兒受體,因此可以迅速從體內循環中清除。因此,當患者治療時為了保持一個恒定的血藥濃度,必須持續使用scFvs。此外,scfv可以作為構建單元來創建bsAbs(圖1C)。
除了增加抗體結構的大小,其他延長抗體半衰期的方法包括與白蛋白融合或與白蛋白結合、與聚乙二醇片段結合以及融合Fc。幾種雙特異性結構與人血清白蛋白融合后,在小鼠模型中增加了半衰期。此外,在雙特異性抗體中添加Fc區域可以避免由于快速清除而需要對小型抗體進行連續給藥。在非人類靈長類動物中,通過融合Fc區域,將不同BiTEs的血清半衰期從6 h延長到44-167 h 。
雙特異性抗體和標準單抗的差別在于雙抗主要作用于兩個不同的靶標。二價結合增加了抗體活性,并可影響其藥效學。雙價抗體可誘導抗體依賴的二聚化,其中一個例子是一種可以阻斷間充質上皮轉化因子(MET)激酶信號。當然,經過改造的單價抗體也可以阻斷MET受體的二聚化和下游信號的激活。
以CD3為靶點的二價抗體會誘導T細胞間的交聯,導致T細胞裂解。但是,靶向CD3的單價抗體在體外卻不會誘導T細胞裂解。為防止接受腎移植的患者出現排斥反應,一種靶向CD3的二價抗體可以發揮耗盡T細胞的作用,但是也會引起嚴重的細胞因子反應。而用于腫瘤治療的能招募免疫細胞參與的bsAbs主要依賴于免疫細胞,發揮抗腫瘤作用因而要避免免疫細胞衰竭,所以這類bsAbs大都用單價抗體和CD3結合。
4.招募免疫細胞
對癌癥免疫治療興趣的日益增長推動了招募免疫細胞的bsAbs的發展。bsAb blinatumomab使免疫細胞與ALL病人的B細胞結合。它能招募表達CD3抗原的T細胞,CD3抗原是T細胞的一般標記物。T細胞通過靶向腫瘤相關抗原(TAA)與腫瘤結合。對于blinatumomab來說,TAA是CD19,CD19是B細胞的一種標志物。一般情況下,TAA應該是針對腫瘤細胞的,使健康組織不受傷害。TAA并不一定在癌癥的發病機制中起作用;其主要作用是在在免疫細胞參與的情況下,在腫瘤細胞膜上提供一個能夠結合的位點。
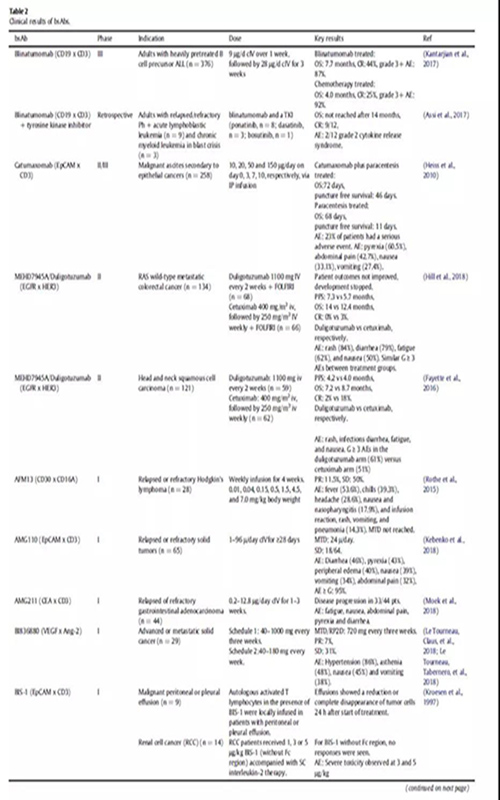
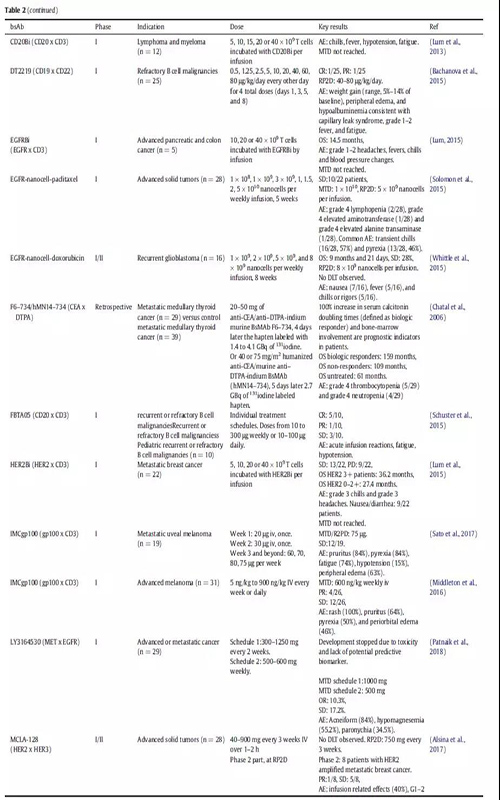
免疫細胞參與bsAbs的應用已經探索了30多年。最近,blinatumomab證實了免疫細胞參與的bsAbs治療血液學惡性腫瘤的潛力。在一項隨機研究中,ALL(heavily pretreated B cell precursor)患者均使用blinatumomab單抗治療,中位生存時間為7.7個月,而化療治療組的中位生存時間為4.0個月(表2)。大多數臨床試驗中的bsAbs都是招募免疫細胞參與的;在ClinicalTrials.gov上報道的57例腫瘤相關的bsAbs中,有38例屬于這種類型(圖3)。
?
4.1 CD3+ T細胞參與的雙特異性抗體
在進行腫瘤臨床試驗的38種通過招募免疫細胞發揮作用的雙特異性抗體中,有36種是通過與T細胞受體CD3結合招募T細胞發揮抗腫瘤作用,在這些雙特異性抗體中有18種針對惡性血液瘤的靶點,16種以實體瘤為靶點。
當T細胞和腫瘤細胞都被bsAb結合時,就形成了一個溶細胞突觸。在這個細胞水解突觸中,T細胞釋放穿孔素和細胞毒性顆粒酶-b,導致靶細胞死亡,這一機理已在體外得到證實,并已通過共聚焦顯微鏡觀察到。在沒有靶細胞的情況下與T細胞結合并不會激活T細胞,這與體外用人外周血單核細胞(PBMCs)和BITE進行的T細胞活化和細胞毒性試驗結果一致。
然而,當表皮生長因子受體(EGFR)陽性和陰性的癌細胞在體外混合并用于建立人類異種移植小鼠模型時,結合CD3和EGFR的BITE也會導致EGFR陰性細胞的死亡。這說明BITE治療也能引起非TAA表達腫瘤細胞的殺傷。
臨床前研究表明,免疫檢查點參與緩解了血液病患者對招募免疫細胞的bsAbs的反應。將以CD33和CD3為靶點的BITE結構雙抗AMG330加入到從患者收集的原發性急性髓系白血病(AML)細胞和PBMCs的共培養中,促進了以AML細胞為主的程序性死亡配體1 (PD-L1)表達的上調(Krupka et al.,2016)。在這些患者樣本中添加抗pd -1和/或抗pd - L1抗體可增強雙抗對AML細胞的殺傷。在食蟹猴中, 采用靶向全長CD3和B細胞譜系標記物FcRH5的bsAb治療多發性骨髓瘤,檢測到血、脾、淋巴結和骨的PD1 + CD8+ T細胞增加及B細胞的消除。將該bsAb在體外與抗PD-L1抗體聯用,可以增加轉染PD-L1編碼質粒的腫瘤細胞的裂解。
在許多具有功能免疫系統的實體瘤小鼠模型中,已經觀察到免疫細胞參與的bsAbs對腫瘤的反應。在這些研究中,選擇了廣泛的TAAs,包括已建立的腫瘤標志物,如癌胚抗原(CEA)、EpCAM、人表皮生長因子受體2 (HER2)和EGFR。然而,關于免疫細胞參與的bsAbs在人類實體瘤中的臨床療效數據卻很少。
一個值得一提的例子是雙特異性抗體IMCgp100,它能結合CD3及黑色素瘤相關抗原糖蛋白-100 (gp100)。IMCgp100的雙特異性抗體ImmTAC以T細胞受體(TCR)而不是抗體的Fab區域作為表面蛋白gp100的靶點。TCRs的使用能夠靶向主要組織相容性復合體分子所表達的細胞內癌蛋白。然而,多克隆T細胞的反應,如cd3參與的bsAbs所產生的反應,被排除在外。針對細胞內wt1蛋白的特異性TCR與靶向CD3的scFv結構的雙特異性抗體,在人類白血病和實體癌的異種移植小鼠模型顯示了療效。
一種稍微不同的方法是使用武裝了T細胞的bsAb。HER2Bi就是一個例子,它是一種bsAb,由靶向HER2和CD3的兩個連接抗體組成。在一期研究中,從患者身上提取T細胞,與bsAb一起培養。然后將T細胞和bsAb重新注入病人體內。由于離體環境中與T細胞結合的條件可控,bsAb的潛在需求更少,副作用的幾率可能會降低。一期臨床研究證實了相對溫和的副作用,并顯示出通常參與抗腫瘤免疫應答的細胞因子增加水平(表2)。
?
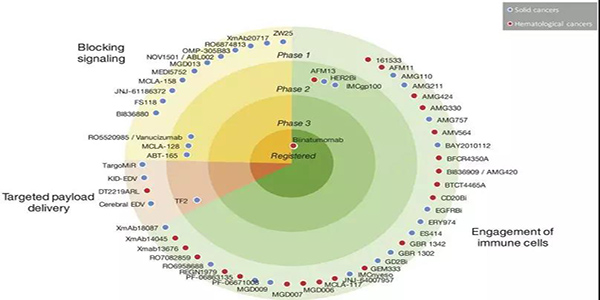
??圖3.正在開發中并在ClinicalTrials.gov網站上注冊了癌癥患者的臨床試驗的BsAbs。BsAbs顯示為點,它們在圖表中的位置表示開發的階段及其作用機制。注冊的bsAbs都顯示在圖表的中心,處于臨床1期的bsAbs顯示在外圍。bsAbs也按照作用機制進行分類:綠色部分表示有免疫細胞的參與的雙抗,紅色部分表示靶向bsAbs,黃色部分表示具有信號阻斷的雙抗。點的顏色表示bsAb是針對實體瘤還是血液瘤。
???4.2招募CD3+ T細胞參與的bsAbs與免疫系統的相互作用
一般來說,動員T細胞的bsAbs破壞其靶細胞是獨立于共刺激信號之外的,如在體外細胞毒性試驗中,在抗cd3×抗cd19 bsAb存在的情況下,人PBMCs誘導人淋巴瘤細胞系的死亡。然而,添加一種共刺激信號,例如白細胞介素-2,可以增強效力,特別是當PBMC與共刺激信號共培養時。同樣,靶向共刺激分子CD137和CD28作為一種聯合治療,可以改善招募免疫細胞的bsAbs的殺傷腫瘤細胞作用。將抗cd137和抗cd20的雙抗與抗cd3和抗cd20的雙抗聯用,在攜帶人類淋巴瘤異種移植瘤的小鼠中顯示出協同作用。然而,單獨使用CD137×CD3 bsAb并不能降低腫瘤的生長。
除了共刺激分子,共抑制分子也被認為阻礙了招募免疫細胞參與的bsAbs的作用。BsAb RO6958688,以CEA和CD3為靶點的2:1 CrossMab結構的雙抗,在與PBMCs聯合移植的小鼠體內增加了T細胞向異種移植結腸癌的浸潤。此外,給予該bsAb可將PD-L1陰性腫瘤轉化為PD-L1陽性腫瘤。同樣的結果也出現在了使用ERY974(一個針對glypican-3和CD3的IgG形式的bsAb)治療移植了轉染人類糖皮質激素-3的肺癌和肝癌細胞的表達人類CD3轉基因小鼠腫瘤模型。在T細胞和一組腫瘤細胞系的體外共培養中,無論初始表達水平如何,靶向CD3和CEA的BiTE均可誘導T細胞上的PD1表達和腫瘤細胞上的PD-L1表達。抗pd1和抗pd - L1抗體的加入增強了BITE抗體的活性。
在體外實驗中,HER2-TBD一種抗her2 x抗cd3的bsAb對轉染PD-L1的HEK293腫瘤細胞的殺傷活性降低。在該研究中,給予該bsAb聯合PD-L1阻斷抗體可恢復bsAb的細胞毒性潛能。接下來,在表達人CD3的轉基因小鼠的同源腫瘤模型中,單獨使用抗her2 x抗CD3 bsAb或聯合抗pd -L1抗體使用,作用于轉染的人her2的CT26腫瘤。聯合治療也能更有效地控制腫瘤生長。結合CD3與TROP-2靶點的Fab(2)-scFv雙抗聯合抗pd1抗體在MDA-MB-231乳腺癌細胞株小鼠體內異種移植瘤模型中顯示了協同作用。
潛在的免疫細胞參與的bsAbs提高了T細胞對實體腫瘤的滲透作用,最新的證據表明抑制PD1 / PD-L1軸可以加強bsAbs的作用,這些結果促使了在I期臨床試驗中評估招募免疫細胞參與的bsAbs聯合檢查點抑制劑的療效,特別是和anti-PD-L1抗體聯用的效果(表3)。
RO6958688是一種針對CEA和CD3的bsAb,聯合抗pdl1抗體atezolizumab治療轉移性結直腸癌臨床結果顯示了聯用具有增強活性的效果。單獨用RO6958688治療的31名患者中有2名患者部分反應,而組合治療組的14名患者中有3名患者部分反應,未發現額外增加的毒性。?
?4.3招募其他免疫受體細胞
除T細胞外,其他效應細胞或免疫細胞亞群也可參與腫瘤細胞的作用。有許多CD3 + T細胞亞型,并不是所有的都有助于抗腫瘤免疫反應。調節性T細胞(Treg)抑制活化的T細胞。42例B細胞ALL患者在blinatumomab單抗治療前外周血中treg的含量與治療反應呈負相關。在體外,blinatumomab激活了抑制效應T細胞細胞毒性的Tregs 。阻止Tregs的活化是CD8+ T細胞與前列腺干細胞抗原串聯結合的scFv結構雙抗背后的理論之一。這種bsAb確實在體外誘導了人前列腺腫瘤細胞株的裂解,但與CD3+ T細胞與人PBMCs和分離的CD8+ T細胞共培養時相比,bsAb的裂解效果較差。
具有作用于T細胞受體CD28與CD20的bsAb在體外的幾種淋巴瘤細胞株和PBMCs共培養體系里面顯示了強勁的殺傷腫瘤細胞的作用。BITE樣結構雙抗RM28以CD28和TAA黑色素瘤相關蛋白聚糖為靶點,作用于黑色素瘤細胞。在2007年完成的一期臨床試驗中,這種bsAb被用于轉移性黑色素瘤患者的腫瘤內治療(NCT00204594),但結果尚不清楚。
BsAbs也被開發用于靶向自然殺傷細胞(NK),這是一種天然免疫系統的強細胞毒性淋巴細胞。一項針對AFM13霍奇金淋巴瘤患者的一期試驗已經完成,這是一種針對CD30和CD16的串聯diabody (TandAb)結構。在該研究中,外周血NK細胞活化,可溶性cd30降低,26例患者中有3例出現部分緩解(Rothe et al.,2015)(表2)。
通過在抗CD33和抗CD16結構之間引入IL-15修飾的bsAb結構(圖1C) 與未修飾的bsAb相比,它具有更好的體外抗腫瘤活性,提高了人NK細胞的存活率。目前計劃對CD33+髓系惡性腫瘤患者進行一項三特異性結構161,533的試驗(表S1)。
?5.?有效載荷遞送
BsAbs也是有效載荷遞送的載體。目前放射免疫治療和抗體-藥物偶聯物等抗體已經進入臨床。在這種方法中,含有同位素或藥物的有效載荷直接與抗體耦合。90Y-ibritumomab tiuxetan注冊用于治療非霍奇金淋巴瘤,曲妥珠單抗抗體偶聯物(ado-trastuzumab emtansine)注冊用于治療轉移性乳腺癌HER2 過表達的患者,brentuximab vedotin登記治療霍奇金淋巴瘤和系統性的間變性大細胞淋巴瘤。它們通過將抗體與TAA結合,將有效載荷直接遞送到腫瘤細胞,帶有有效載荷的抗體,與TAA結合,然后被內在化,有效載荷在細胞內釋放并發揮作用。
使用bsAb可以實現新的目標定位方法。與抗體直接偶聯不同,與TAA和有效載荷有親和力的bsAb可以在注射前與有效載荷結合。通過首先注射對TAA和有效載荷具有親和力的bsAb,然后注入有效負載,也可以實現預定目標的傳遞。用于向腫瘤遞送有效載荷的預靶向技術可能潛在地規避健康組織長時間暴露于有效載荷,從而減輕毒性和不良反應。連接有效載荷和bsAb是通過將bsAb的一只手臂指向有效載荷的半抗原來實現的。半抗原是一種本身不具有免疫原性的分子,但可以作為抗原并與抗體結合。
1993年發表了第一篇報告使用bsAb遞送有效載荷的臨床試驗的論文。目前,有5種bsAbs正在進行臨床試驗,其中4種針對實體腫瘤。BsAb TF2,有三個Fab片段,其中兩個靶向CEA和一個靶向有效載荷,目前最快推進到2期臨床(圖3)。 ?
5.1放射性有效載荷的預定投放
表達CEA的髓樣甲狀腺癌患者注射bsAb TF2,靶向CEA和有效載荷。24小時后,施用有效負載,即用111銦標記的小肽。在施用該小肽后24小時觀察腫瘤與組織中有效負載的比率大于1:20,顯示用bsAbs預靶向的可行性。理論上,由于其體積小,未綁定的有效載荷將被迅速清除,減小了對非目標組織的損傷。
當有效載荷為治療性放射金屬時,半抗原可作為放射金屬的螯合劑。另一種選擇是使用兩個半抗原創建一個大的雙價半抗原,有利于同兩個腫瘤結合的bsAbs,這將使腫瘤的結合更加穩定。該系統被稱為親和增強系統,已用于臨床研究(表2)。
對于用于放射免疫療法的預靶向遞送釔-90,將CD38和DOTA-釔復合物具有親和力的bsAb與通過鏈霉抗生物素蛋白-?生物素鍵結合放射性金屬的抗體進行比較。在異種移植的非霍奇金淋巴瘤或多發性骨髓瘤小鼠中,bsAb方法顯示出優于鏈霉親和素生物素方法的抗腫瘤效果(Green et al.,2018)。也可以通過連接有效載荷和抗體的替代方法實現預靶向。這些包括鏈霉親和素-?生物素,寡核苷酸或click化學,如四嗪和反式環辛烯之間的環加成反應。然而,bsAbs方法是迄今為止唯一在臨床中進行測試的方法(表2)。
5.2?其他有效載荷的遞送
bsAbs通過結合螯合劑半抗原,在動物模型中探索了bsAb(如阿霉素)預先靶向遞送其他有毒有效載荷。在這些研究中,螯合劑被加載放射性同位素锝-99,以驗證目標特異性結合。其他半抗原,如地高辛,也可以結合到有效載荷上,用于藥物傳遞。幾種有效載荷,如阿霉素和與地高辛結合的熒光染料Cy5,在人類異種移植小鼠模型中顯示出特定的靶向性。
一種直接靶向的方法,即在給藥前將bsAb和有效載荷孵育的方法正在進行臨床實驗。在這種方法中,有效載荷被封裝在一個細菌衍生的納米細胞中,稱為engeneic delivery vehicle (EDV),bsAbs是通過Fc區域連接在一起的兩個抗體。有效載荷可以是化療藥物,如阿霉素或紫杉醇,但也可以是microRNA。三個測試EDVs的試驗結果已經公開,一期臨床數據顯示安全狀況尚可接受。
bsAb DT2219與有效載荷直接偶聯,并且靶向CD22和CD19以增強特定的遞送能力。有效載荷為白喉毒素,經CD19和/或CD22內化后進入胞質。該bsAb已在難治性B細胞惡性腫瘤患者中進行研究,25例患者中有1例出現完全反應和1例出現部分反應(表2)。
6.?信號阻斷
用聯合療法靶向癌癥的多個表位或受體是一種流行的方法,許多抗體治療的聯合方法正在臨床試驗中進行評估。
抗pd -1抗體nivolumab與抗ctla4抗體ipilimumab聯合治療轉移性黑色素瘤已獲FDA和EMA批準。最近,這種組合也被FDA批準用于治療晚期腎細胞癌。另一種稍微不同的聯合治療是使用帕妥珠單抗和曲妥珠單抗的多表位方法,兩者都以HER2為靶點,但在不同的表位上。它已被批準作為轉移性her2陽性腫瘤的聯合治療。
從理論上講,兩種抗體的靶點可以合并成一個單一的bsAb,可以產生各種各樣的好處。這種藥物的特異性可以通過癌癥受體的共同定位來增強,從而將健康組織的靶向毒性降到最低。此外,可以通過靶向一個抗原的不同表位來提高結合親和力。這種bsAb的潛在缺點是,它將自己限制在一個固定的抗原組合,而單抗可以自由組合,可以不同順序給藥或個性化劑量給藥。根據ClinicalTrials.gov網站的數據,目前正在臨床試驗中研究14種bsAbs,它們可以阻斷重要的腫瘤信號傳導。
6.1腫瘤細胞表面受體
由于不同信號之間相互作用,普通bsAbs主要干擾兩個信號途徑,如ErbB家族成員EGFR、HER2和HER3。
BsAbs MM-111、JNJ-61186372和MEHD7945A是針對這些目標中的一個或多個。盡管它們的半衰期都很長(表1),但是它們使用不同的結構來實現這一點。
有趣的是,針對EGFR和HER3的bsAb MEHD7945A比抗EGFR抗體西妥昔單抗或EGFR 激酶抑制劑厄洛替尼更有效,在人非小細胞肺癌和頭頸部鱗癌的異種移植模型中克服了西妥昔單抗和厄洛替尼耐藥。這很可能是由于阻斷了ErbB家族成員的信號通路的交互作用。然而,MEHD7945A對轉移性結直腸癌和頭頸部鱗狀細胞的2期試驗中沒有發現MEHD7945A優于西妥昔單抗的益處。因此,這個bsAb的開發已經停止(表2)。
正在研究的其他靶標有死亡受體,如CD95,或參與溶酶體內化的受體,如CD63。以CD20和cd95為靶點的bsAb比不同的抗CD20抗體變體更能抑制人類異種移植瘤小鼠的腫瘤生長。為了提高抗體藥物偶聯物,我們設計了一種載藥bsAb,它除了結合HER2外,還結合受體CD63。熒光共聚焦顯微鏡顯示,能夠誘導其內化,改善了her2陽性異種移植小鼠模型的腫瘤抑制作用。
CD47-SIRPα交互,也稱之為“別吃我信號”,通過在巨噬細胞上表達的SIRPα抑制表達CD47的細胞的吞噬作用和該靶點過表達在許多固體和血液腫瘤細胞上。這種相互作用也可能被bsAbs破壞。在與Raji腫瘤細胞異種移植的小鼠中,靶向CD20和CD47的IgG-scFv bsAb延長了小鼠的生存期,靶向CD19和CD47的IgG-like bsAb根除了腫瘤,而抗cd47、抗cd20或抗cd19抗體是無效的。
靶向SIRPα在用Burkitt淋巴瘤異種移植的小鼠中不誘導腫瘤消退,盡管結合anti-CD20抗體美羅華導致協同效應和作用于SIRPα和CD70的bsAb減緩腫瘤的生長。然而,bsAb與抗SIRPα抗體聯合抗CD70抗體產生的腫瘤生長減少相同。 ?
6.2免疫受體
隨著免疫檢查點抑制劑的建立及其在腫瘤治療中的結合,bsAbs正在探索作為這些現有治療的補充或改進。四價雙親和重定向(DART)結構的MGD013以兩價同時靶向淋巴細胞活化基因3 (lag3)和pd -1;它將在晚期實體瘤患者的臨床試驗中得到驗證。在體外,MGD013較單一治療或聯合治療引起T細胞因子釋放增加,提示T細胞活化增加。
MEDI5752是一種結合PD-1和CTLA-4抑制腫瘤浸潤淋巴細胞的抗體。這將在晚期實體腫瘤患者的臨床試驗中進行測試。
IgG樣雙抗FS118通過其Fab片段靶向PD-L1,通過其Fc區域靶向lag3,也阻斷了兩條通路。FS118的小鼠抗體,靶向小鼠Lag-3和PD-L1,通過增加CD8:Tregs比值,劑量依賴性的誘導了其抗腫瘤活性,改變了免疫浸潤淋巴細胞的組成。該結構正在晚期癌癥患者的臨床試驗中進行測試(表S1)。
6.3抑制血管生成
除了抑制細胞膜上的靶標,腫瘤環境本身也可以成為靶點。Crossmab雙抗vanucizumab通過作用于血管生成素-2 (ang2)和血管內皮生長因子- a (VEGF-A)來抑制血管生成。bsAb OMP-305B83靶向δ樣配體4和VEGF。在該結構中,兩種bsAb都是帶有Fc的,因為長的半衰期對于效的因子消耗是至關重要的。
Vanucizumab可抑制多種基因的患者來源的異種移植瘤模型小鼠的腫瘤生長和轉移。它還增加了腫瘤內免疫細胞的激活,導致內皮細胞上調PD-L1的表達(在多個同源小鼠模型中已經驗證)。在該方法中,在vanucizumab中添加抗pd -1抗體治療可提高存活率,為進一步合理評價該bsAb聯合免疫治療提供了依據(表3)。
6.4增加特異性
bsAb RO6874813是一個2:2的Crossmab,它采用了一種不同的方法。它與腫瘤細胞上激活TNF相關凋亡誘導配體受體之一的死亡受體(DR) 5和腫瘤相關成纖維細胞上的纖維母細胞活化蛋白(FAP)具有親和力。與之前用抗體激活腫瘤細胞上的DR5的嘗試不同,這種bsAb通過對癌癥相關成纖維細胞的親和力增強了對腫瘤的特異性。在體外和人移植成纖維細胞與不同的癌癥或病人來源的肉瘤組合的小鼠模型中,這種bsAb的有效性取決于與癌癥相關的成纖維細胞的存在。在體內模型中,bsAb比抗dr5治療更有效地抑制腫瘤生長。
7.?剩余的挑戰
blinatumomab和emicizumab的批準刺激了bsAbs進入臨床試驗(圖4)。在采用小分子量雙抗治療患者時,需要持續給藥,如blinatumomab,以保持恒定的血藥濃度。解決這一不足的一種方法是通過添加Fc區域延長bsAbs的半衰期。
目前,兩種流行的小型bsAb平臺BiTE和DART平臺,在臨床試驗中都有Fc區域擴展版本(圖1C)。針對DLL3和CD3的AMG757是BITE-Fc;MGD007和MGD009分別靶向糖蛋白A33和CD3、B7-H3和CD3,都是為DART-Fc結構。所有這些bsAbs都針對實體腫瘤。MGD007最近完成了復發或難治性轉移性結直腸癌患者的一期臨床試驗(NCT02248805),目前研究結果尚未發表。然而,MGD007的研究設計說明了更長的半衰期的優勢;使用每周和三周的治療方案,而靶向CD123和CD3的DART分子MGD006通過持續靜脈輸注給AML患者(NCT02152956)。越來越多進入臨床試驗的新型bsAbs具有Fc區域(圖4)。
此外,blinatumomab通過逐步給藥來減輕毒性。這種結構的嚴重毒性是由一種叫做細胞因子釋放綜合征的全身細胞因子釋放引起的,常見于T細胞參與治療中。除了逐步給藥,皮質類固醇也用于減少細胞因子釋放綜合征。
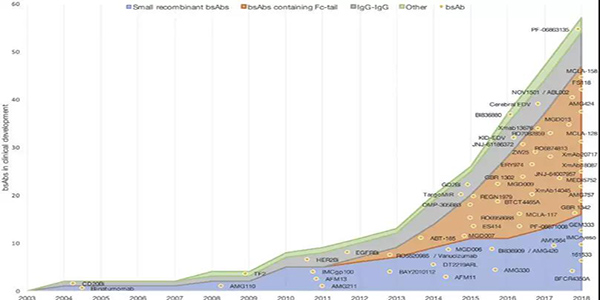
圖4正在開發中并在ClinicalTrials.gov網站上注冊針對癌癥患者開展臨床試驗 的BsAbs。線條顯示每年使用的結構數量,bsAbs顯示為點。他們在圖表中的位置顯示了所使用的結構和他們第一次臨床試驗的開始日期。?
最近,通過細胞因子水平檢測,在人類B細胞淋巴瘤的小鼠模型中,使用抗CD20抗體預處理后,給予CD20-和cd3靶向Crossmab bsAb可降低毒性。在該研究設計中,預處理使外周血和繼發性淋巴器官中的B細胞減少,從而防止了它們的意外激活,避免了招募免疫細胞的bsAb釋放細胞因子。
此外,最近對同源小鼠腫瘤模型的研究表明,具有不同CD3親和力的her2靶向的bsAbs的分布存在差異。CD3的高親和力降低了全身暴露,并向淋巴組織轉移攝取)。另一項研究表明,參與CD3和c型凝集素樣分子-1的bsAb的副作用依賴于CD3的親和力:在食蟹猴實驗中高親和力變異導致細胞因子釋放水平增高。
這些發現強調了對像bsAbs這樣的新結構進行廣泛的藥代動力學研究的必要性,例如通過分子成像。雙特異性抗體結構的設計是一個挑戰,因為該藥物的生物分布是由該結構的兩個部分以及該結構的所有其他藥效學特性決定的。雖然有許多方法來測量新藥的藥代動力學,但分子成像是唯一的非侵入性方法。
分子成像研究可用于建立雙特異性結構部分藥代動力學的預測模型,并制定最佳的給藥策略。這對于所有尚未在臨床試驗中評估的不同結構尤其重要。
分子成像用于藥物動力學研究的一個例子是一種用于正電子發射斷層掃描的鋯89標記的AMG211示蹤劑。AMG211是一種針對CEA和CD3的BiTE。在對晚期胃腸道腺癌患者的I期試驗中,使用這種方法對轉移瘤進行了成像。患者體內和患者之間存在異質性腫瘤攝取,淋巴組織中存在cd3特異性攝取。 ?
8.?結論 ?
評估這些藥物的臨床試驗證明,人們對bsAbs作為治療癌癥的一種方法非常感興趣。bsAb目前用于臨床實踐,但沒有一種正在進行癌癥治療的三期臨床試驗。大多數被評估的bsAbs具有相同的作用機制:免疫細胞與腫瘤細胞的結合。在遞送有效載荷方面,使用bsAbs的熱情似乎由于諸如click化學等簡單的共軛方法的出現而有所減弱。
臨床前研究表明,免疫細胞參與的bsAbs與抗pd1、抗pd - l1等免疫調節劑聯合使用后,其抗腫瘤作用增強。最初的臨床結果證實了這一點,但還需要更多的數據。將進入臨床試驗的不同而新穎的bsAbs結構也為使用分子成像揭示其體內行為提供了強有力的論據。近年來,bsAb已成為一種多用途的工具,但除了blinatumomab,它還沒有導致臨床突破。然而,由于越來越容易生產以及雙特異性抗體獨特的作用機制,bsAbs可能成為一個有價值的抗腫瘤軍火庫的補充。
參考文獻:
A review of bispecific antibodies and antibody constructs in oncology and clinical challenges
如涉及知識產權,請與我司聯系!??
|


